
Трехмерная модель процессов эндогенного голографического управления развитием пространственной структуры биосистем
|
© И.В.Прангишвили, П.П.Гаряев, Г.Г.Тертышный, А.В.Мологин, Е.А.Леонова, Э.Р.Мулдашев. Ивери Варламович Прангишвили – д-р техн. наук,
акад. АН Грузии, директор ИПУ РАН; Тел. (095) 335-18-06, 253-21-70, 466-33-89; e-mail gariaev@aha.ru РЕЗЮМЕ В предыдущей нашей статье [1], основанной на работе [2], мы предложили плоскую 2-мерную транспорантно-голографическую модель пространственного развития биосистем (биоморфогенеза). В настоящей работе предложена 3-мерная модификация этой модели. Как и в [1], допустим существование в генетико-информационных
процессах поляризационно-волнового кода, что развивает ранее предложенную
[3, 4] модель волнового самоуправления в биосистемах. Нам представляется
вероятным наличие в пространстве оптически активного хромосомного
жидкокристаллического континуума многоклеточных организмов некоторого
гено-биознакового дискретного динамического диапазона частот и углов
поляризации векторов Умова-Пойтинга когерентных световых излучений
хромосом. Относительно реальности таких излучений in vivo не приходится
сомневаться [10], что подтверждается нашей работой по прямой лазерной
накачке ДНК и хромосом in vitro [6]. В биосистеме при таких поляризационно-
и частотно модулированных пространством организма излучениях происходит
резонансный информационно-волновой обмен и кодовое оптическое узнавание
волновых сигналов, несущих как генетическую, так и метаболическую
информацию. Нарушения в механизмах такого узнавания, например, при
зашумлении объемного голографического считывания и распознавания
образов, ведут к информационно-хаотическому состоянию волновых систем
управления организмом, и прежде всего, в генетическом аппарате.
В этом плане важно понять, как голографическое кодирование на уровне
хромосомного континуума биосистем работает в объемном варианте.
Вероятно, основные закономерности оптики [7] должны выполняться
и объяснять сверхслабые знаковые световые процессы, происходящие
в организмах. Это один из этапов волнового управления биосистемой
на фотонном уровне. Однако, волновая регуляция метаболизмом организмом
этим не ограничивается, но включает в себя также и управление поляризациями
радиоволн, связанными с модуляционными поляризациями фотонных полей
при квантово-нелокальном переходе фотоны Учитывая многослойное строение голографических транспарантов, известных в технике, и многослойные жидкокристаллические упаковки хромосом [5], рассматриваемые нами как аналоги транспарантов, можно представить многократное и избыточное наложение одних и тех же восстановленных образов, каждый из которых является одновременно и голограммой-транспарантом и просто транспарантом для случая его повреждения. Причем, при существенной степени повреждения транспарант реконструирует изображение с меньшим разрешением, чем тот, который не был поврежден. При суммарном нарастании объемной массы в процессе развития из яйцеклетки биосистема приобретает новые калибровочные (разметочные) градиенты света при его дифракции на ее клеточно-тканевых (в том числе хромосомных) объемных структурах-аналогах голограмм (биоголограмм). При этом длины реконструирующих волн могут существенно возрастать, достигая в упругих волнах сантиметрового и дециметрового диапазонов. Градиенты этих волн управляют геометрическими параметрами растущих и развивающихся структур эмбриона. Для классификации голограмм используют параметр
где n – средний коэффициент преломления хромосомного
транспаранта; d – толщина транспаранта; Принято считать голограмму толстой (объемной) при
значении Q Рассмотрим процесс амплитудной и фазовой записи как тонких, так и толстых биоголограмм. Пространственное изменение амплитудного пропускания света через клеточно-тканевые биослои можно выразить в виде
Это выражение характеризует амплитудную биоголограмму. Амплитудное пропускание в оптике определяют обычно как отношение интенсивностей прошедшего к интенсивности падающего лучей когерентного света:
Обе величины связаны соотношением: T = Линейная запись может быть описана выражением
Производная ( d Пространственная модуляция интенсивности света в интерференционном поле I = A в результате воздействия на регистрирующий транспарант фиксируется как пространственная модуляция амплитудного пропускания развивающейся послойно биоткани. В соответствии с выражением (1) можно
записать Величины Вещества, входящие в состав организма и составляющие
его, различным образом взаимодействуют со светом (записывают информацию),
что влияет на амплитуду проходящего через биоткани световой волны,
а от фазовой структуры биоткани зависит фазовая задержка в этой
биоструктуре. Такая фазовая задержка определяется разностью хода
Ф = (2 Сопоставляя формулу (3) с
известным уравнением Фазовую структуризацию поля световой волны биосистема осуществляет как путем пространственной модуляции коэффициента преломления локальной биоткани, т.е. ее вещественного состава, так и путем модуляции длины оптического пути в биоткани, т.е. посредством изменения толщины нарастающего транспаранта (фрагмента биосистемы). Кроме того, фазовая структура может задаваться путем сложного построения поверхностного рельефа биомембран, эндоплазматического ретикулума, границ раздела жидкокристаллических фаз жидких кристаллов хромосом и т.д. На такой поверхности свет, дифрагируя, создает сложный узор спекл-картины, которая также может являться информационно-голографической системой управления ростом, развитием и поддержанием биосистемы в заданных рамках геометрической структуры. Анализируя фазовую структуру биоголографических объемных транспарантов, ограничимся случаем линейного отклика формируемого биоматериала, т.е. линейной зависимостью фазы световой волны от количества внешнего и внутреннего освещения. При этом внешнее освещение носит естественный характер (например, солнечное освещение), а внутреннее формируется в биосистемах за счет эндогенных лазероподобных процессов в молекулах ДНК хромосом и в белках [4, 6].
При этом амплитудное пропускание биотканей является
комплексной величиной, т.е.: Абсолютная величина пропускания рассматриваемых сред равна единице и изменяется только фазовый член. Подставив уравнение (5) в (6), получим для случая небольших изменений пропускания света биотканями
где показательную функцию exp [ i (d при необходимости заменим первыми двумя членами разложения в ряд. Рассмотрим процесс реконструкции волнового фронта
на объемных биоголограммах. Этот процесс происходит посредством
дифракции на трехмерной дифракции Фраунгофера плоской волны на плоском
фрагменте объемной биорешетки, в качестве которых могут выступать
жидкокристаллические среды биосистемы. Их можно рассматривать как
совокупность равноудаленных изофазных плоскостей. При падении света
на фрагмент объемной биоголограммы под углом U В данном случае опускаются квадратичные члены в показателе
экспоненты; коэффициент К содержит в себе все константы;
Приведенное уравнение описывает дифракцию Фраунгофера на плоском фрагменте объемной биоголограммы. Для трехмерной решетки предположим, что это уравнение описывает дифракцию на элементарном слое, который можно считать биорешеткой из-за того, что тонкий слой производит такое же дифракционное рассеивание как это происходит на обычных голографических тонких решетках. толщиной dz’, а результирующая амплитуда определяется суммой волн, дифрагированных на отдельных элементарных транспарантах. Подобный подход является решением задачи в первом приближении. В этом случае данное уравнение необходимо дополнить дифракционным интегралом по сечению биофрагмента голограммы, перпендикулярному ее поверхности. В результате получим U Х exp[ik'(m где l – направляющие косинусы. В случае, если реконструирующая и дифрагированная
биоволны образуют с осью z' углы l m Функция Интенсивность интерференционного поля, образованного референтной и объектной волнами, как обычно определяется следующим образом [7]: I = ( u Разность фаз Ф Ф В последнем уравнении тригонометрические функции можно заменить направляющими косинусами: l m Теперь уравнение (11) можно записать в виде Ф Таким же образом можно выразить направляющие косинусы
интерференционных полей, изофазные биоповерхности которых образуют
с осью z' угол L = cos В соответствии с углом наклона плоскостей, определяемым как tg для угла tg Подставив это уравнение в (13), получим Ф где
Величина Подставив (16) в (10), определим интенсивность интерференционного поля : I = A В случае линейной записи амплитудное пропускание фрагмента биосистемы в соответствии с уравнением (1) будет равно
Последнее выражение преобразуем к виду
где W U
+exp[-i
Это выражение можно представить в виде суммы трех интегралов: U
+K Интегралы можно вычислить, используя соотношение
Для удобства обозначим sinBA/(BA) = sinc(BA). Тогда получим уравнение (22) в виде: U
Для реконструкции объемной биоголограммы, так же
как и для случая плоской голограммы, конечное выражение для амплитуды
дифрагированной волны распадается на три части. Проанализируем третий
член этого уравнения. Его значение определяется функцией sinc Максимум этой функции имеет место при
т.е. при условии равенства углов , т.е. когда
Равенство углов означает, что восстановленная биоволна тождественна реконструирующей биоволне, т.е. – волне нулевого порядка дифракции. Первый и второй члены уравнения описывают дифрагированные волны плюс первого и минус первого порядков, т.е. основное и вторичное голографические изображения. Рассмотрим это на примере первого члена. Он достигает максимума при условии :
Оба уравнения выражают так называемые условия Брэгга для дифракции на объемной биологической решетке. В случае плоской биологической решетки для первого порядка дифракции выполняется соотношение sin где Уравнение (25) можно преобразовать к виду m где Учитывая, что полосы биорешетки являются линиями пересечения интерференционных плоскостей с плоскостью z = 0, и выражая расстояние между этими полосами через расстояние между интерференционными плоскостями, преобразуем уравнение (26) к виду
что соответствует второму уравнению (24). Первое условие Брэгга идентично уравнению биорешетки, а первый дифракционный максимум объемной биорешетки идентичен первому максимуму плоской биорешетки, которая является элементарным слоем анализируемой объемной биорешетки. Возвращаясь к первому уравнению (24), воспользуемся законом зеркального отражения
В соответствии с этим же законом зеркального отражения следует, что :
Используем условие дифракции (27), тогда окончательно получим
Формула (30) идентична уравнению (24) и выражает
условие Брэгга, согласно которому вектор реконструирующей волны
должен совпадать с таким направлением, чтобы зеркально отраженная
интерференционно-построенная биоплоскостями волна совпадала с дифрагированной
волной. Все эти рассуждения аналогичны и в отношении второго уравнения
(24). Отличие состоит в том, что этот член содержит
Поскольку
откуда следует:
Применяя известные формулы тригонометрии, находим: В таком случае аргументы функций sinc равны:
где верхний знак относится к первичному изображению фрагмента биоструктуры, а нижний - к вторичному. При умножении на мнимую единицу второго уравнения (33) и сложении с его первым уравнением, получим комплексное выражение, которое представим в виде суммы показательных функций:
Уравнение (34) будет равно нулю при максимальной
интенсивности дифрагированной волны на биофрагменте голограммы.
При векторном представлении всех членов последнего уравнения сумма
их векторов будет равна нулю. Если длины биоволн реконструирующей,
предметной и референтной совпадают, а это значит, что
а соответствующие векторы образуют ромб. Рассмотрим выражение с верхним знаком при следующих трех условиях: №1 №2 №3 Затем рассмотрим это же выражение с нижним знаком при других условиях: №1 – 1 №2 – 2 №3 – 3 Выражения №1 и № 2 – 2 соответствуют основному и вторичному изображениям. При выполнении условий №2 и №1 –1 реконструкция имеет место только в случае дифракции на равномерной регулярной объемной биологической решетке, образованной двумя плоскими волнами. Качественного изображения при реконструкции сложной световой волны, не получится, поскольку в данном случае не выполняется условие Брэгга одновременно для всех составляющих пространственно-частотного спектра этой волны. Отсюда следует, что объемные биоголограммы как регистрируются, так и реконструируются сравнительно простыми монохроматическими световыми фронтами и “работают” они последовательно друг за другом на сравнительно близком расстоянии. При этом не будет происходить искажений, вызванных рассеянием когерентного света на этих биоструктурах. Условия №3 и №3 – 3 соответствуют случаю отсутствия интерференционной картины и создают при этом некогерентный фон. Это условие хорошо соответствует условию применимости фонового принципа для биосистем, которым они пользуются в полной мере [4, 8]. Итак, биосистемы, вероятно, используют как плоские (тонкие) биоголограммы, так и объемные (толстые), которые совместно, дополняя друг друга, выполняют стратегически важную роль при построении трехмерного пространства живого организма. Это не исключает, однако, и другие формы биоморфогенеза, связанные, к примеру, с использованием тексто-подобных структур ДНК и РНК или с формообразовательными функциями солитонных доменных стенок, структурирующих организм [3, 4, 9], а также поляризационно-радиоволновые процессы самоорганизации [11]. Убедительным подтверждением принципов волновой самоорганизации биосистем служит первое в мире успешное приживление имплантированных слепому человеку донорских тканей с восстановлением зрения [12]. В основе идеологии таких хирургических операций и пост операционных регенеративных процессов лежат наши исследования [1, 3, 4, 6, 8, 11], включая настоящую работу. Литература 1. И.В.Прангишвили, П.П.Гаряев, Г.Г.Тертышный, Е.А.Леонова, А.В.Мологин, М.Р.Гарбер, 2000, Генетические структуры как источник и приемник голографической информации. Датчики и Системы, №2, с.3-8. 2. Д. Бом, “Квантовая теория” пер. с англ. Л.А. Шубиной под ред. С.В. Вонсовского, изд.”НАУКА” , М., 1965 г. стр.187. 3. Гаряев П.П. Волновой геном. М. Изд. Общ. польза. 279с. 1994г. 4. Гаряев П.П. Волновой генетический код. М. Издатцентр. 107с. 1997г. 5. Livolant F. Condensed forms of DNA in vivo and in vitro // Physics A. 1991.V.176. P.117-137. 6. А.М.Агальцов, П.П.Гаряев, В.С.Горелик, И.А.Рахматуллаев, В.А.Щеглов, 1996, Двухфотонно-возбуждаемая люминесценция в генетических структурах. Квантовая электроника, v.23, N2, с.181-184. 7. М. Милер, Голография, Л., “МАШИНОСТРОЕНИЕ”, 1979г. 8. Прангишвили И.В., Ануашвили А.Н., Маклаков В.В. Закономерности проявления подвижного объекта / Сб. трудов ИПУ РАН. М. Вып. 1. С.7-10. (1993). 9. Петухов С.В., 1999, Биосолитоны - тайна живого вещества. Основы солитонной биологии. М. Изд. “ГП Кимрская типография”. 288с. 10. Biophotonics and coherent systems. Proceedings of the 2-nd A.Gurwitsch Conference and Additional Contributions. Moscow University Press-2000. Ed. L.Beloussov et al. M. 445p. 11. И.В.Прангишвили, П.П.Гаряев, Г.Г.Тертышный, В.В.Максименко, А.В.Мологин, Е.А.Леонова, Э.Р.Мулдашев, 2000, Спектроскопия радиоволновых излучений локализованных фотонов: выход на квантово-нелокальные биоинформационные процессы. Датчики и Системы, №9(18), с.2-13. 12. Мулдашев Э.Р., Комбинированная трансплантация глаза. Министерство здравоохранения Российской Федерации, Всероссийский Центр Глазной и Пластической Хирургии. “Аллоплант”, 2000. |




 (22)
(22) (23)
(23)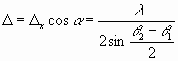 имеем:
имеем: 
 ,
поскольку
,
поскольку